Gallimimus bullatus (lat. "similar a una gallina con bulla") es la única especie conocida del género extinto Gallimimus de dinosaurio terópodo ornitomímido, que vivió a finales del período Cretácico, hace aproximadamente 70 millones de años en el Maastrichtiense, en lo que hoy es Asia.
Fue hallado en la Formación Nemegt, en Mongolia. Debido a su constitución ligera y sus largas patas traseras, este dinosaurio estaba adaptado para la carrera. Sin duda fue esa su gran braza para escapar de los depredadores. Su aspecto recuerda al de un avestruz dado su largo cuello y su pico sin dientes; no obstante, no hay constancia de que tuviera alas o plumas. Su larga cola le servía para mantener el equilibrio al avanzar.
Descripción[]
Gallimimus es el miembro más grande conocido de la familia Ornithomimidae. El holotipo adulto, espécimen IGM 100/11, tenía aproximadamente 6 metros de largo y 1,9 metros de altura en la cadera. Su cráneo tenía 330 milímetros de largo y el fémur, hueso del muslo, tenía 660 milímetros. Habría pesado unos 440 kilogramos. En comparación, un espécimen juvenil, ZPAL MgD-I / 94 tenía aproximadamente 2,15 metros de largo, 0,79 metros de altura en la cadera y pesaba alrededor de 26 kilogramos. Sobre la base de los fósiles de los parientes Ornithomimus, se sabe que los ornitomímidos tenían plumas y los adultos portaban estructuras en forma de alas, como lo demuestra la presencia de perillas en el hueso del cúbito de la parte inferior del brazo, protuberancias que indican dónde se habrían adherido las plumas.
Gallimimus era en apariencia similar a los actuales avestruces, cabeza pequeña, grandes ojos, cuello largo, miembros anteriores pequeños, fuertes patas para correr y una larga cola. Un carácter de diagnóstico de Gallimimus son sus manos cortas en comparación al húmero, cuando se lo compara con otros ornitomímidos. Sus patas delanteras eran relativamente cortas comparadas con las traseras, y estaban rematados con tres garras en cada mano. A pesar de estar afiladas, no está claro que tuvieran mucha utilidad. Algunos expertos han dado utilidad a las garras del Gallimimus aventurando que pudieron servirle para escarbar en la tierra y conseguir así huevos, raíces y tubérculos con los que alimentarse. Además de esto, en la forma mongol los arcos neurales dorsales de las vértebras sacras no se funden totalmente incluso en especímenes maduros, en contraste con esos observados en Ornithomimus y Struthiomimus. La cola era utilizada como contrapeso. Los ojos estaban situados a los lados de s
u cabeza, significando que no poseía la visión binocular. Como la mayoría de los pájaros modernos, tenía huesos huecos. Gallimimus tenía un número de adaptaciones que sugieren buena capacidad para correr, miembros largos, con una tibia y metatarso largos y los dedos del pie cortos. Un pico fosilizado presente en un cráneo de Gallimimus estaba bordeado de un pico que se ha interpretado como parte de un mecanismo de filtrado como los de los patos. Sin embargo, picos similares se ven en tortugas de mar herbívoras y los ornitomímidos eran relativamente comunes en ambientes con estaciones secas, donde no había alimentación para filtrar. Parece más probable que Gallimimus fuese un omnívoro, usando su pico en las plantas y la captura de pequeños animales. Sobre el Gallimimus se ha especulado sobre su dieta, sugiriendo que pudo haberse alimentado de insectos y de pequeños gusanos y pudo complementar su dieta insectívora con algunas plantas y huevos.
Cráneo[]
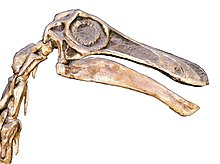

{kind=link}
Cráneo y cuello reconstruidos, NHM
La cabeza de Gallimimus era muy pequeña y ligera en comparación con la columna vertebral. Debido a la longitud de su hocico, el cráneo era largo comparado con otros ornitomímidos y el hocico tenía un perfil superior ligeramente inclinado. Esto era menos distintivo en especímenes juveniles. El perfil lateral del hocico se diferenció de otros ornitomímidos al no estrecharse hacia su mitad frontal y el margen frontal inferior de la premaxila en la parte frontal de la mandíbula superior se elevó hacia arriba, en lugar de ser horizontal. Visto desde arriba, el hocico era casi espatulado, en forma de cuchara, ancho y redondeado en la punta , en forma de U, mientras que era agudo o en forma de V en los ornitomímidos norteamericanos. Las cuencas de los ojos eran grandes y estaban orientadas hacia los lados, como en otros ornitomímidos. La región temporal al lado del cráneo detrás de los ojos era profunda, y la fenestra infratemporal, la abertura inferior detrás de la órbita, era casi triangular y más pequeña que la del Struthiomimus. Tenía cicatrices musculares profundas en la parte posterior del techo del cráneo, a lo largo del hueso parietal. El paraesfenoide, un hueso en la parte inferior de la base del cráneo, tenía paredes delgadas, huecas y formaba una estructura bulbosa en forma de pera. La estructura tenía un surco poco profundo que se abría en la base del cráneo. Las narinas internas, aberturas internas para el pasaje nasal, eran grandes y estaban colocadas muy atrás en el paladar, debido a la presencia de un extenso paladar secundario , que era común a los ornitomímidos.


{kind=link}
Cráneo de un ejemplar juvenil de Gallimimus.
La delicada mandíbula inferior, formada por huesos delgados, era delgada y poco profunda en la parte delantera, profundizándose hacia la parte posterior. La parte frontal de la mandíbula tenía forma de pala, lo que resultaba en un espacio entre las puntas de las mandíbulas cuando se cerraba. La forma similar a una pala era similar a la de la gaviota común y el pico inferior puede haber tenido una forma similar a la de esta ave. El proceso retroarticular en la parte posterior de la mandíbula,;donde se unían los músculos de la mandíbula, estaba bien desarrollado y consistía principalmente en el hueso angular. El surangular era el hueso más grande de la mandíbula inferior, como es habitual en los terópodos. La fenestra mandibular, una abertura orientada hacia los lados en la mandíbula inferior, era alargada y comparativamente pequeña. La mandíbula inferior no tuvo un proceso coronoideo o un hueso supradentario, cuya falta es una característica común de los terópodos con pico, ornitomimosaurios, ovirraptorosaurianos, tericinosaurianos y aves, pero inusual entre los terópodos en general. La forma y las proporciones del cráneo cambiaron significativamente durante el crecimiento. La parte posterior del cráneo y las órbitas disminuyeron de tamaño, mientras que el hocico se hizo relativamente más largo, se producen cambios similares en los cocodrilos modernos . El cráneo también fue proporcionalmente más grande en los ejemplares más jóvenes. Las mandíbulas de Gallimimus estaban desdentadas, sin dientes y la parte frontal se habría cubierto de un pico córneo en vida. El pico puede haber cubierto un área más pequeña que en los parientes de América del Norte, debido a la falta foramenes nutritivos en el maxilar. El lado interno del pico tenía estructuras columnares pequeñas, apretadas y espaciadas uniformemente, se discute su naturtaleza exacta, que eran más largas en la parte delantera y acortadas hacia la parte posterior.
Esqueleto postcraneal[]
Gallimimus tenía 64–66 vértebras en su columna vertebral , menos que otros ornitomímidos. Los centros o cuerpos de las vértebras eran platycoelas, con una superficie frontal plana y una superficie trasera cóncava, a excepción de las seis primeras vértebras caudales, donde la superficie trasera también era plana, y aquellas al final de la cola que eran anfiplatas, con ambas superficies planas. Muchos de los centros tenían foramenes, aberturas que también se han llamado pleurocoelos y, por lo tanto, probablemente eran neumáticas, con sus cámaras huecas invadidas por sacos de aire. El cuello consistía en 10 vértebras cervicales, todas largas y anchas, excepto el atlas, la primera vértebra que se conecta con la parte posterior del cráneo. El atlas difería del de otros ornitomímidos en que la superficie de su intercentro estaba inclinada hacia abajo hacia la parte posterior, en lugar de ser cóncava y hacia arriba. El cuello parece haber sido proporcionalmente más largo en relación con el tronco que en otros ornitomímidos. El cuello se dividió en dos secciones distintas, las vértebras cervicales en la parte delantera tenían centros que eran casi triangulares en vista lateral y se estrechaban hacia la parte posterior, así como arcos neurales bajos y cigapófisis anchas y anchas los procesosque articulado entre las vértebras, las vértebras cervicales en la parte posterior tenían centros con forma de carrete que se volvieron gradualmente más altos y cigapófisis largas y delgadas. La resistencia neumática aquí era pequeña y ovalada y las espinas neurales que sobresalían de los centros formaban crestas largas, bajas y afiladas, excepto en las vértebras cervicales más posteriores.
La espalda de Gallimimus tenía 13 vértebras dorsales, con centros en forma de carrete que eran cortos, pero tendían a ser más profundos y más largos hacia la espalda. Sus procesos transversales, procesos articulados con las costillas, aumentan ligeramente de longitud hacia la espalda. Los dos primeros centros dorsales tenían cavidades neumáticas profundas, mientras que el resto solo tenía depresiones poco profundas, y las espinas neurales eran prominentes y tenían una forma algo triangular o rectangular. El sacro, las vértebras fusionadas entre los huesos de la pelvis, consistía en cinco vértebras sacras que eran aproximadamente iguales en longitud. Los centros aquí eran en forma de carrete, aplanados hacia los lados y tenían fosas que parecían haber continuado como foramen profundo en algunos especímenes. Las espinas neurales aquí eran rectangulares, anchas y más altas que las de las vértebras dorsales. Eran superiores o iguales en altura al margen superior de la lamina iliaca y estaban separados, mientras que en otros ornitomímidos se fusionaron. La cola tenía 36–39 vértebras caudales, con los centros de los de la parte frontal en forma de carrete, mientras que las de la parte posterior eran casi triangulares y alargados. Las espinas neurales aquí eran altas y planas, pero disminuían hacia atrás, donde se hacían como una cresta. El único signo de neumaticidad en la cola fueron las fosas profundas entre los espinas neurales y el proceso transversal de las dos primeras vértebras caudales. Todas las vértebras en frente del sacro tenían costillas a excepción del atlas y la última vértebra dorsal. Las costillas en el cuello se fusionaron con las vértebras solo en adultos.
La escápula era corta y curvada, delgada en la parte delantera y gruesa en la parte posterior. Estaba conectado relativamente débilmente con el coracoides, que era grande y profundo de arriba abajo. En general, las extremidades anteriores no diferían mucho de las de otros ornitomímidos, todos los cuales eran comparativamente débiles. El húmero, el hueso del brazo superior, que tenía una sección transversal casi circular, era largo y retorcido. La cresta deltopectoral en la parte frontal superior del húmero era comparativamente pequeña y, por lo tanto, proporcionaba poca superficie para unir los músculos de la parte superior del brazo. El cúbito era delgado, largo y débilmente curvado, con un eje casi triangular. El olecranon, la proyección desde el codo, fue prominente en adultos, pero no bien desarrollada en juveniles. El radio, el otro hueso en el brazo inferior, era largo y delgado con un extremo superior más expandido en comparación con el inferior. La mano fue proporcionalmente corta en comparación con el de otros ornitomimosaurianos, ya que tenía la relación más larga entre la mano y húmero de cualquier miembro del grupo, pero por lo demás era similar en su estructura. Tenía tres dedos, que se desarrollaron de manera similar; el primero' el "pulgar", fue el más fuerte, el tercero fue el más débil y el segundo fue el más largo. Los unguales, los huesos de garras, eran fuertes, algo curvados, el del primer dedo era más curvo y comprimidos lateralmente con un surco profundo en cada lado. Los unguales se desarrollaron de manera similar, aunque el tercero fue ligeramente más pequeño. Las extremidades anteriores parecen haberse vuelto proporcionalmente más largas durante el crecimiento.El cociente del antebrazo con el brazo alcanza el 66% en Gallimimus, mientras que en las formas americanas varia del 70% a incluso el 91% en Dromiceiomimus. Otra diferencias con sus parientes de Norteamérica se ven en las comparciones de las longitudes de los huesos. En Gallimimus el cociente de las vértebras dorsales con las caudales del 85% es mayor que en otros géneros, en Struthiomimus, por ejemplo, este cociente es el aproximadamente 76%.
El pubis era largo y delgado, terminando en una bota púbica que se expandió hacia delante y hacia atrás, una característica general de los ornitomimosaurianos. Las extremidades posteriores diferían poco de las de otros ornitomímidos, y eran proporcionalmente más largas que en otros terópodos. El fémur era casi recto, largo y delgado con un eje aplanado lateralmente. La tibia era recta, larga, con dos cóndilos bien desarrollados, los extremo redondeado de un hueso, en el extremo superior y un extremo inferior aplanado. El peroné de la pierna inferior era plano, delgado y ancho en el extremo superior, estrechándose hacia el extremo inferior. La mitad inferior del tercer metatarsiano era ancho cuando se veía al final, cubriendo en parte los dos metatarsianos adyacentes a cada lado, pero se estrechó abruptamente en la mitad de la longitud, encajándose entre esos huesos y desapareciendo, una estructura de pellizco arctometatarsal. El tercer dedo del pie fue proporcionalmente más corto en relación con la extremidad que en otros ornitomímidos. Al igual que en otros ornitomímidos, el pie no tenía hallux, el primer dedo de la mayoría de los demás terópodos. Los unguals de los dedos de los pies eran planos en sus lados inferiores, los dos exteriores declinaron ligeramente hacia afuera desde sus dígitos. La longitud proporcional de los huesos en las extremidades posteriores cambió muy poco con el crecimiento. Las sacrales en Gallimimus también se distinguen por su gran alargamiento concerniente a la longitud del fémur.
Descubrimiento e investigación[]
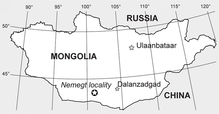
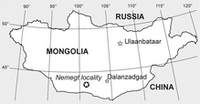
{kind=link}
La localidad de Nemegt (✪) en la cuenca de Nemegt, en el sur de Mongolia, donde se han encontrado algunos especímenes de Gallimimus.
Entre 1963 y 1965, la Academia de Ciencias de Polonia y la Academia de Ciencias de Mongolia organizaron las expediciones paleontológicas polaco-mongoles al desierto de Gobi en Mongolia. Entre los restos de dinosaurios descubiertos en los lechos de arena de la cuenca Nemegt se encontraban numerosos ornitomímidos en diferentes etapas de crecimiento de las localidades dev Nemegt, Tsaagan Khushuu, Altan Ula IV y Naran Bulak. Se recogieron tres esqueletos parcialmente completos, dos con cráneos, así como muchos restos fragmentarios. La paleontóloga Zofia Kielan-Jaworowska descubrió el esqueleto más grande, que luego se convertiría en el holotipo de Gallimimus bullatus en Tsaagan Khushuu en 1964, se conservó acostado sobre su espalda y se encontró el cráneo debajo de su pelvis. También se encontró un pequeño espécimen en Tsaagan Khushuu el mismo año, y otro pequeño espécimen se encontró en la localidad de Nemegt. Un pequeño esqueleto sin extremidades anteriores fue descubierto en 1967 por la expedición paleontológica de Mongolia en Bugeen Tsav fuera de la Cuenca Nemegt. Los fósiles se alojaron en la Academia de Ciencias de Mongolia, Polonia y la URSS. Las expediciones polaco-mongoles fueron notables por ser dirigidas por mujeres, algunas de las cuales fueron de las primeras mujeres en nombrar nuevos dinosaurios. Los fósiles descubiertos en estas expediciones arrojaron nueva luz sobre el intercambio de fauna entre Asia y América del Norte durante el Cretácico. Algunos de los esqueletos se exhibieron en Varsovia en 1968, montados en posturas altas y semi-erectas, que fueron aceptadas en ese momento, aunque hoy en día se prefieren más posturas horizontales.


{kind=link}
Esqueleto de Gallimimus
En 1972, los paleontólogos Rinchen Barsbold, Halszka Osmólska, y Ewa Roniewicz nombraron al género. La única especie conocida y tipo Gallimimus bullatus, utilizando el esqueleto recolectado más grande, el espécimen IGM 100/11, de Tsaagan Khushuu, anteriormente conocido como GINo.DPS 100/11 y MPD 100 / 11, como el holotipo. El nombre genérico se deriva del latín gallus, "pollo" y del griego mimos, "imitador", en referencia a la parte frontal de las vértebras del cuello que se asemejaban a las de los galliformes. El nombre específico se deriva del latín bulla , una cápsula de oro usada por los romanos jóvenes alrededor del cuello, en referencia a la cápsula bulbosa en el parasfenoide en la base del cráneo del dinosaurio. Tal característica no había sido descrita por otros reptiles en ese momento, y era considerada inusual. El holotipo consiste en un esqueleto casi completo con un hocico distorsionado, mandíbula inferior incompleta, series vertebrales, pelvis, así como algunos huesos faltantes de manos y pies. Los otros esqueletos parcialmente completos eran juveniles. ZPAL MgD-I/1, de Tsaagan Khushuu, tiene un cráneo aplastado con una punta faltante, vértebras dañadas, costillas fragmentadas, faja pectoral y extremidades anteriores, y una extremidad posterior izquierda incompleta. ZPAL MgD-I/94 de la localidad de Nemegt, carece de cráneo, atlas, punta de la cola, faja pectoral y extremidades anteriores, mientras que el espécimen más pequeño. IGM 100/10 de Bugeen Tsav, carece de faja pectoral, extremidades anteriores y varias vértebras y costillas. Osmólska y sus colegas enumeraron 25 especímenes conocidos en total, 9 de los cuales solo estaban representados por huesos individuales.
En el momento en que recibió su nombre, los fósiles de Gallimimus representaba el material ornitomímido más completo y mejor conservado que se haya descubierto y el género sigue siendo uno de los miembros más conocidos del grupo. Los ornitomímidos se conocían anteriormente principalmente en América del Norte, siendo Archaeornithomimus el único miembro conocido anteriormente de Asia, aunque sin cráneo. Desde los primeros descubrimientos, más expediciones internacionales lideradas por mongoles han encontrado más especímenes. Tres de los esqueletos de Gallimimus, incluido el holotipo, más tarde se convirtieron en parte de una exhibición itinerante de fósiles de dinosaurios de Mongolia, que recorrieron varios países.
La caza furtiva de fósiles se ha convertido en un grave problema en Mongolia en el siglo XXI y varios especímenes de Gallimimus han sido saqueados. En 2017, Hang-Jae Lee y sus colegas informaron sobre un rastro fósil descubierto en 2009 asociado con un pie de Gallimimus, el espécimen MPC-D100F/17. El resto del esqueleto parecía haber sido eliminado previamente por cazadores furtivos, junto con varios otros especímenes de Gallimimus, como lo indican los pozos de excavación vacíos, la basura y los huesos rotos dispersos en la cantera. Es inusual encontrar huellas estrechamente asociadas con fósiles corporales. Algunas de las pistas son consistentes con los pies de ornitomímidos, mientras que otras pertenecen a diferentes dinosaurios.
En 1988, el paleontólogo Gregory S. Paul concluyó que los cráneos de los ornitomímidos eran más similares entre sí de lo que se pensaba anteriormente y movieron a la mayoría de las especies al mismo género, Ornithomimus, lo que dio como resultado la nueva combinación Ornithomimus bullatus. En 2010, en su lugar usó la combinación Struthiomimus bullatus. Las especies involucradas generalmente han sido mantenidas en géneros separados por otros escritores. En una revista de 1988 se informó sobre una vértebra de ornitomímido de Japón llamada "Sanchusaurus", pero fue asignada a Gallimimus sp. por el paleontólogo Dong Zhiming y sus colegas en 1990.
Barsbold se refirió informalmente a un esqueleto casi completo, "IGM 100/14' como "Gallimimus mongoliensis", pero como se diferencia de Gallimimus en algunos detalles, Yoshitsugu Kobayashi y Barsbold propusieron en 2006 que probablemente Pertenece a un género diferente. Un reciente análisis del esqueleto casi completo de "G. mongoliensis" concluyó que no pertenece a Gallimimus pero representa un nuevo, todavía no nombrado género de ornitomímido.
En el año 2000, el paleontólogo Philip J. Currie propuso que Anserimimus, que solo se conoce de un esqueleto de Mongolia, era un sinónimo menor de Gallimimus, pero esto fue descartado por Kobayashi y Barsbold, quienes señalaron varias diferencias entre los dos. Barsbold notó alguna variación morfológica entre los nuevos especímenes de Gallimimus, aunque esto nunca ha sido publicado.
Nuevos fósiles descubiertos por Peter J. Makovicky en el Desierto de Gobi en el 2001, indican que Gallimimus poseía un pico que filtraba de los charcos animales diminutos y agua a través de unas ranuras o lamelas que tienen en el pico y la lengua como los flamencos actuales.
Clasificación[]
Osmólska y sus colegas asignaron a Gallimimus a la familia Ornithomimidae en 1972, con el norteamericano Struthiomimus como el pariente más cercano, al tiempo que lamentaban el hecho de que la comparación entre los taxones era difícil porque otros ornitomímidos conocidos en ese momento estaban mal conservados o inadecuadamente descritos. En 1975, Kielan-Jaworowska declaró que aunque muchos dinosaurios de Asia se ubicaban en las mismas familias que los parientes norteamericanos, esta categoría de clasificación tendía a ser más inclusiva de lo que se usaba para las aves modernas. Destacó que mientras Gallimimus tenía un pico redondeado, similar a un ganso o un pato, los ornitomímidos norteamericanos tenían picos puntiagudos, una diferencia que de otro modo llevaría a los taxónomos a colocar aves modernas en diferentes familias. En 1976, Barsbold colocó Ornithomimidae en el nuevo grupo Ornithomimosauria, los "dinosaurios avestruz". En 2003, Kobayashi y Jun-Chang Lü encontraron que Anserimimus era el taxón hermano de Gallimimus , ambos formando un clado derivado o "avanzado" con géneros norteamericanos, lo que fue confirmado por Kobayashi y Barsbold en 2006.
Los ornitomimosaurios pertenecían al clado Maniraptoriformes de los terópodos celurosaurianos, que también incluye a las aves modernas. Los ornitomimosaurianos tempranos tenían dientes, que se perdían en más miembros derivados del grupo. En 2004, Makovicky, Kobayashi y Currie sugirieron que la mayor parte de la historia evolutiva temprana de los ornitomimosaurianos tuvo lugar en Asia, donde se descubrieron la mayoría de los géneros, incluidos los taxones más basales o "primitivos", aunque reconocieron que La presencia del basal Pelecanimimus en Europa presenta una complicación en la clasificación. El grupo debe haberse dispersado una o dos veces desde Asia a Norteamérica a través de Beringia para dar cuenta de los géneros del Cretácico Tardío que se encuentran allí. Como se ve en algunos otros grupos de dinosaurios, los ornitomimosaurianos se restringieron en gran medida a Asia y América del Norte después de que Europa se separara de Asia por el estrecho de Turgai.
En 1994, el paleontólogo Thomas R. Holtz agrupó ornitomimosaurianos y troodóntidos en un clado, basándose en características compartidas, como la presencia de una cápsula bulbosa en el parasfenoide. Llamó al clado Bullatosauria, basándose en el nombre específico de Gallimimus bullatus, que también se refería a la cápsula. En 1998, Holtz descubrió que los troodóntontidos eran maniraptoranos basales, lo que significa que todos los miembros de ese clado caerían dentro de la Bullatosauria, que por lo tanto se convertiría en un sinónimo menor de Maniraptoriformes, y desde entonces el clado ha quedado fuera de uso.
Paleobiología[]
Las vértebras cervicales de Gallimimus indican que sostuvo su cuello oblicuamente, inclinando hacia arriba en un ángulo de 35 grados. Osmólska y sus colegas encontraron que las manos de Gallimimus no eran prensiles, capaces de agarrar, y que el pulgar no era oponible. También sugirieron que los brazos eran débiles en comparación con, por ejemplo, los del ornitomimosauriano Deinocheirus. Esto estaba de acuerdo con las interpretaciones de la biología de los ornitomímidoservicios realizadas por el paleontólogo Dale Russell desde principios de 1972, incluidas las que hubieran sido animales corredores, aunque menos ágiles que las grandes y modernas aves terrestres y habrían utilizado su velocidad para escapar de los depredadores. Russell también sugirió que tenían un buen sentido de visión e inteligencia comparable al de las aves ratites modernas. Dado que sus depredadores pueden haber tenido visión a color, sugirió que habría influido en su coloración, lo que tal vez resultara en camuflaje. En 1982, el paleontólogo Richard A. Thulborn estimó que Gallimimus podría haber corrido a velocidades de 42-56 km/h. Encontró que los ornitímidos no habrían sido tan rápidos como los avestruces, que pueden alcanzar los 70-80 km/h, en parte debido a que sus brazos y colas aumentan su peso.
En 1988, Paul sugirió que los globos oculares de los ornitomímidos se aplanaban y tenían una movilidad mínima dentro de las cuencas, lo que requería el movimiento de la cabeza para ver los objetos. Dado que sus ojos estaban más orientados hacia los lados que en otros terópodos parecidos a aves, su visión binocular habría sido más limitada, lo cual es una adaptación en algunos animales que mejora su capacidad para ver a los depredadores detrás de ellos. Paul consideró que las colas relativamente cortas, que reducían el peso y los halux faltantes de ornitomímidos eran adaptaciones para la velocidad. Él sugirió que podrían haberse defendido a sí mismos picoteando y pateando, pero habrían dependido principalmente de su velocidad para escapar. En 2015, Akinobu Watanabe y sus colegas encontraron que junto con Deinocheirus y Archaeornithomimus, Gallimimus tenía el esqueleto más neumatizado entre los ornitomimosaurios. Se piensa que la neumatización es ventajosa para el vuelo en aves modernas, pero su función en dinosaurios no aviares no se conoce con certeza. Se ha propuesto que la neumatización se usó para reducir la masa de los huesos grandes, que se relacionó con un alto metabolismo, el equilibrio durante la locomoción o se usó para la termorregulación.
En 2017, Lee y sus colegas sugirieron varias posibles circunstancias tafonómicas, los cambios durante la descomposición y la fosilización para explicar cómo el pie de Gallimimus descubierto en 2009 se asoció con una vía. La pista se conserva en piedra arenisca, mientras que el pie se conserva en piedra arenisca , extendiéndose 20 centímetros debajo de la capa con la pistas. Es posible que el fósil represente a un animal que murió en su camino, pero la profundidad del pie en el lodo puede ser demasiado superficial para que se haya quedado empantanada. También puede haber sido matado por una inundación, después de lo cual fue enterrado en un estanque. Sin embargo, las capas de lodo y arena no indican inundaciones pero probablemente un ambiente seco y los sedimentos desorganizados alrededor del fósil indica que el animal estaba vivo cuando llegó a la zona. Por lo tanto, los autores sugirieron que las huellas se habían realizado durante un período prolongado de tiempo y período de secado, y que probablemente ninguna de ellas fue producida por el individuo que poseía el pie. El animal puede haber caminado por el suelo de un estanque, atravesando la capa de sedimento con las huellas mientras estaba empapado de la lluvia o el agua contenida. El animal puede haber muerto en esta posición por sed, hambre u otra razón, y luego se deposita lodo en la arena, cubriendo y preservando las huellas y el cadáver. El pie pudo haberse apretado y desarticulado al descomponerse, lo que hizo que los tendones se flexionaran, y luego fue pisado por dinosaurios pesados. El área puede haber sido un solo lecho óseo, basado en el posible número de especímenes capturados, que representan una mortalidad masiva de Gallimimus, tal vez debido a una sequía o hambruna. El hecho de que los animales parecen haber muerto al mismo tiempo, los pozos de excavación vacíos eran estratigráficamente idénticos, puede indicar que Gallimimus era gregario, lo que también se ha sugerido para otros ornitomímidos.
Ontogenia[]
Un estudio realizado en 1987 por los biólogos R. Pawlicki y P. Bolechała mostró diferencias relacionadas con la edad en el contenido de calcio y fósforo,:componentes importantes en la formación de hueso, de los especímenes de Gallimimus. Encontraron que la proporción era más alta en animales jóvenes y de mediana edad, disminuyendo con la edad. En 1991, informaron que los huesos de individuos viejos contenían las mayores cantidades de plomo y hierro, mientras que los de los animales más jóvenes eran más bajos. Un estudio de la histología ósea de varios dinosaurios en 2000, por los biólogos John M. Rensberger y Mahito Watabe, reveló que los canalículos, canales que conectan las células óseas y el colágeno, los haces de fibras, de Gallimimus y otros ornitomímidos eran más similares a los de las aves que a los mamíferos, a diferencia de los dinosaurios ornitisquios, que eran más similares a los mamíferos. Estas diferencias pueden haber estado relacionadas con el proceso y la velocidad a la que se formó el hueso. En 2012, la paleontóloga Darla K. Zelenitsky y sus colegas concluyeron que, dado que los ornitomimosaurianos adultos tenían estructuras en forma de alas en sus brazos, mientras que los juveniles no, como lo demuestran los especímenes de Ornithomimus, estas estructuras eran originalmente características sexuales secundarias , que podrían se han utilizado para el comportamiento reproductivo, como el cortejo, la visualización y la crianza.
Estrategias de alimentación[]
Osmólska y sus colegas señalaron que la parte frontal del cuello de Gallimimus habría sido muy móvil y la parte trasera era más rígida, los arcos neurales en las vértebras de esa región eran similares a los de los gallos y otros galliformes, lo que indica hábitos de alimentación similares. Encontraron el pico de Gallimimus similar al de un pato o ganso y se habría alimentado de presas pequeñas y vivas que se tragó entero. La movilidad del cuello hubiera sido útil para ubicar a la presa en el suelo, ya que los ojos estaban colocados a los lados del cráneo. Asumieron que todos los ornitomímidos tenían hábitos de alimentación similares y señalaron que Russel había comparado los picos de los ornitomímidos con los de las aves insectívoras. Osmólska y sus colegas sugirieron que Gallimimus fue capaz de cinesis craneal, debido a la conexión aparentemente suelta entre algunos de los huesos en la parte posterior del cráneo, una característica que permite que los huesos individuales del cráneo se muevan entre sí. También propusieron que no usara sus extremidades anteriores para llevar comida a la boca, sino para rastrillar o cavar en el suelo para acceder a la comida. Las manos de Gallimimus pueden haber sido más débiles que, por ejemplo, las de Struthiomimus, que en su lugar pueden haber usado sus manos para enganchar y agarrar, según un artículo de 1985 de los paleontólogos Elizabeth L. Nicholls y Anthony P. Russell.
En 1988, Paul no estuvo de acuerdo en que los ornitomímidos eran omnívoros que comían animales pequeños, huevos y plantas, como se había sugerido anteriormente. Señaló que los avestruces y los emúes son principalmente animales de pastoreo y ramoreadores y que los cráneos de los ornitomímidos eran más similares a los de los extintos moas, que eran lo suficientemente fuertes como para masticar las ramitas, como lo demuestra su contenido intestinal. Además, sugirió que los ornitomímidos estaban bien adaptados para alimentarse de plantas resistentes y que habrían usado sus manos para llevar las ramas al alcance de sus mandíbulas. El paleontólogo Jørn Hurum sugirió en 2001 que debido a su estructura de mandíbula similar, Gallimimus puede haber tenido una dieta omnívora, oportunista, como las gaviotas. También observó que la unión intramandibular apretada evitaría cualquier movimiento entre las partes delantera y trasera de la mandíbula inferior.
En 2001, los paleontólogos Mark A. Norell, Makovicky y Currie publicaron un cráneo de Gallimimus, IGM 100/1133 y un cráneo de Ornithomimus que conservaban estructuras de tejidos blandos en el pico. El lado interno del pico de Gallimimus tenía estructuras columnares que los autores encontraron similares a las laminillas en los picos de las aves anseriformes, que las utilizan para manipular alimentos, filtrar sedimentos, separar los alimentos de otros materiales y para cortar plantas durante el pastoreo. Encontraron al pato cuchara, que se alimenta de plantas, moluscos, ostracodos y foraminíferos, como el anseriforme moderno con estructuras más similares en anatomía a las de Gallimimus. Los autores notaron que los ornitomímidos probablemente no usaban sus picos para cazar animales grandes y eran abundantes en ambientes mésicos, mientras que eran más raros en ambientes más áridos, lo que sugiere que pueden haber dependido de fuentes de alimentos acuáticos. Si esta interpretación es correcta, Gallimimus habría sido uno de los animales filtradores terrestre más grandes conocidos.
En 2005, el paleontólogo Paul Barrett señaló que las estructuras similares a lamelas de Gallimimus no parecían ser cerdas flexibles como las de las aves que se alimentan por filtración, ya que no hay indicios de que estas estructuras se superpongan o se colapsen, sino que fueron más similar a las crestas verticales delgadas y regularmente espaciadas en los picos de las tortugas y dinosaurios hadrosáuridos. En estos animales, se cree que dichas crestas están asociadas con dietas herbívoras, utilizadas para moler vegetación resistente. Barrett sugirió que las crestas en el pico de Gallimimus representaban un molde natural de la superficie interna del pico, lo que indicaba que el animal era un herbívoro que se alimentaba de material con alto contenido de fibra. El descubrimiento de muchos gastrolitos, piedras de molleja, en algunos ornitomímidos indican la presencia de un molino gástrico y por lo tanto, apuntan hacia una dieta herbívora, ya que se utilizan para moler alimentos de animales que carecen del aparato de masticación necesario. Barrett también calculó que unos 440 kilogramos Gallimimus habría necesitado entre 0,07 y 3,34 kilogramos de alimento por día, dependiendo de si se tenía un metabolismo endotérmico o uno ectotérmico, una ingesta que no era factible si se trataba de un filtrador. También descubrió que los ornitomímidos eran abundantes no solo en formaciones que representaban ambientes mésicos, sino también en ambientes áridos donde no habría suficiente agua para mantener una dieta basada en la alimentación por filtración. En 2007, el paleontólogo Espen M. Knutsen escribió que la forma de pico de los ornitomímidos, en comparación con los de las aves modernas, era consistente con un omnívoro o un herbívoro de vegetales altos en fibra.
Paleoecología[]
Gallimimus es conocido de la formación Nemegt en el desierto de Gobi en el sur de Mongolia. Esta formación geológica nunca se ha fechado radiométricamente, pero la fauna presente en el registro fósil indica que probablemente se depositó durante la etapa Maastrichtiense temprana, al final del Cretácico Superior hace unos 70 millones de años. Los sedimentos de la localidad tipo de Gallimimus, Tsaagan Khushuu constan de limos, limolitas, lutitas, arenas, así como menos frecuentes capas delgadas de areniscas. Las facies rocosas de la formación Nemegt sugieren la presencia de canales de ríos, marismas, lagos poco profundos y planicies aluviales en un entorno similar al delta del Okavango de la actual Botsuana. Los grandes canales fluviales y los depósitos de suelo son evidencia de un clima significativamente más húmedo que el de las formaciones más antiguas de Barun Goyot y Djadochta. Aunque los depósitos de caliche indican que se produjeron sequías periódicas. Los huesos fósiles de la cuenca Nemegt, incluyendo Gallimimus, son más radioactivos que los fósiles de las áreas circundantes, posiblemente porque el uranio se acumula en los huesos, transportado allí mediante la filtración de agua subterránea.
Los ríos Nemegt, donde vivía Gallimimus, eran el hogar de una gran variedad de organismos. En esta región se han descubierto fósiles de moluscos ocasionales , así como una variedad de otros animales acuáticos como peces, tortugas y crocodilomorfos, incluido el Shamosuchus. Los fósiles de mamíferos son raros en la formación Nemegt, pero se han encontrado muchas aves, incluida la Enantiornithes Gurilynia, el Hesperornithes Judinornis y Teviornis, un posible anseriforme. Los dinosaurios herbívoros descubiertos en la formación Nemegt incluyen anquilosáuridos como Tarchia, el paquicefalosauriano Prenocephale, grandes hadrosáuridos como Saurolophus y Barsboldia y saurópodos como Nemegtosaurus y Opisthocoelicaudia. Los terópodos depredadores que vivían junto a Gallimimus incluyen tiranosauroides como Tarbosaurus, Alioramus y Bagaraatan, y troodóntidos como Borogovia, Tochisaurus y Zanabazar. Los terópodos herbívoros u omnívoros incluyen tericinosaurianos, como Therizinosaurus, así como oviraptorosaurianos, como Elmisaurus, Nemegtomaia y Rinchenia. También se encuentran otros ornitomimosaurios, incluidos Anserimimus y Deinocheirus, pero Gallimimus es el miembro más común del grupo en el Nemegt
En la franquicia de Jurassic Park[]
Aparece en los dos primeros filmes de Jurassic Park: en el primero es perseguida una manada de ellos por el Tyrannosaurus rex, siendo uno de los ejemplares una comida ligera para el enorme depredador, y en la segunda película vemos ejemplares siendo capturados por cazadores, luego se lo ve fugazmente mientras huye del campamento cuando Nick Van Owen abre las jaulas y libera a los animales. Aparece también nombrado como una de las especies de la Isla Sorna en la novela de The Lost World Jurassic Park, y tambien aparece en muchos juegos y comics. En la difunta pagina Jurassic Park Institute, podemos ver ademas de a Gallimimus, a un sinonimo menor llamado Sanchusaurus, esta especie hoy por hoy es invalida.
Historia[]
Este dinosaurio fue clonado a principios de la década de 1990 por International Genetic Technologies, Inc. en Isla Sorna después de algunos años de investigación a partir de 1983. Se obtuvo ADN antiguo de ámbar mongol del Cretácico con inclusiones de mosquitos y otros organismos bebedores de sangre, complementado con ADN de anfibios modernos para reemplazar segmentos descompuestos del genoma de los dinosaurios. En junio de 1993, InGen había creado hasta la versión 2.035 de este animal. Actualmente está presente en el noroeste del Pacífico después de ser liberado en la naturaleza a mediados de 2018.
Descripción[]

{kind=link}
Con un crecimiento de entre 20 y 26 pies (6,1 a 7,9 metros) de largo, los Gallimimus de InGen son más grandes que los conocidos por los fósiles, pero no se sabe si esto es el resultado de la ingeniería genética o de la vida en cautiverio. Los animales criados más recientemente han promediado ser un pie más altos que los creados inicialmente por InGen, lo que sugiere que las buenas condiciones de vida pueden ser la explicación. Pueden alcanzar 6,2 pies (1,9 metros) de altura en las caderas, y los adultos pesan entre 500 y 1000 libras (226,8 y 453,6 kilogramos), siendo 970 libras (439,98 kilogramos) el peso promedio sugerido por los fósiles.
Gallimimus es notablemente diferente de la mayoría de los otros terópodos no aviares, ya que está adaptado a un estilo de vida de forrajeo en lugar de cazador. Si bien la mayoría de los terópodos son carnívoros depredadores, Gallimimus es omnívoro y se alimenta principalmente de plantas y animales pequeños. Este dinosaurio carece de dientes y, en cambio, tiene un pico queratinoso similar al de un pájaro; tiene laminillas en forma de dientes similares a las de los gansos. Estas laminillas le ayudan a agarrar los alimentos en la boca, especialmente los alimentos más resbaladizos como los gusanos y los anfibios. También pueden ayudarlo a cortar la materia vegetal. Su lengua rosada es estrecha y puntiaguda, mientras que el pico es aplanado y rosado. La forma de U de la punta de su hocico la diferencia de las ornitomimidas norteamericanas, que tienen puntas de hocico en forma de V.
Tiene ojos grandes que miran hacia los lados con pupilas redondas; los iris son estrechos y oscuros, a menudo indistinguibles, pero las escleróticas son claramente amarillas. Los ojos son planos y no móviles dentro de la cuenca. Combinado con la visión binocular reducida, esto significa que Gallimimus necesita girar toda la cabeza para cambiar la dirección en la que mira. Su visión no es completamente estereoscópica, pero se parece más a la de un animal de presa típico que a la de un cazador.
La cabeza de este dinosaurio está encaramada sobre un cuello largo y delgado, que es algo (aunque no extremadamente) flexible y le da un buen punto de vista desde el cual ver su entorno. Al levantar la cabeza en alto, puede inspeccionar el área cercana para estar atento a los depredadores. Normalmente, el cuello y la cabeza se mantienen en un ángulo de treinta y cinco grados en comparación con el cuerpo. Los hombros y los brazos son comparativamente débiles, siendo los brazos más cortos que los de otros ornitomimidos. Sus manos tienen tres dedos, cada uno de los cuales termina en una garra larga; a pesar de su tamaño, son ineficaces como armas porque carecen de poderosos músculos en los brazos. Su primer dedo es el más fuerte, mientras que el tercero es el más débil; su dedo medio es el más largo. Como ocurre con la mayoría de los terópodos InGen, las muñecas del Gallimimus está en pronación, a diferencia del animal original.
Si bien sus brazos son débiles, no se puede decir lo mismo de sus poderosas piernas. Los huesos de sus piernas son largos, delgados y rectos; sus caderas y patas son musculosas y capaces de impulsar a este animal hacia adelante a velocidades de treinta a treinta y cinco millas por hora (48,3 a 56,3 kilómetros por hora). Algunas fuentes como Jurassic World: The Game sugieren que, en un sprint muerto, la velocidad de este dinosaurio podría alcanzar un máximo de sesenta millas por hora (96,6 kilómetros por hora), aunque probablemente no podría mantener esa velocidad durante mucho tiempo debido al peso de su cola y brazos. Para ayudarlo a agarrarse en el suelo, sus tres dedos terminan en garras curvas. Como todos los ornitomimidos pero a diferencia de otros tipos de terópodos, carece de espolón. El cuerpo de Gallimimuses delgado y atlético, con huesos huecos como un pájaro, y generalmente está construido para ser ágil y de pies rápidos. Su cola es corta y anodina, y sirve en gran medida para mantener el equilibrio mientras corre.
Los guiones anteriores de Jurassic Park, y algunos materiales suplementarios restantes de 1993, describen huevos y crías de Gallimimus en la naturaleza (en la película final, este papel fue desempeñado por Velociraptor). Esto significaría que los Gallimimus de InGen pueden cambiar de sexo de hembra a macho, un proceso llamado protoginia, y reproducirse. Sin embargo, nunca se ha confirmado realmente. Si este es el caso, su genoma probablemente incluye estructuras tomadas de la rana común, Hyperolius viridiflavus.
En la prehistoria, este dinosaurio habría tenido plumas, particularmente formando estructuras en forma de alas en los brazos, haciéndolo parecerse aún más a un avestruz moderno. Sin embargo, debido a complicaciones en la ingeniería genética, la mayoría de los genomas de terópodos de InGen incluyen un alelo nulo donde normalmente se produciría el gen para el desarrollo de las plumas y, en su lugar, desarrollarían piel escamosa. Gallimimus se encuentra entre estos casos, mostrando pequeñas escamas en forma de guijarros que cubren la mayor parte de su cuerpo y escamas rectangulares más grandes en los dígitos similares a muchas aves modernas. Estas son probablemente las únicas regiones de su cuerpo donde su piel puede parecerse a la de su antepasado, ya que las aves tienen escamas similares en sus patas. Las fuentes no están de acuerdo sobre si InGen tenía la intención de crear especímenes emplumados, resolviendo errores genómicos, o si los especímenes escamosos se desempeñaron mejor con las audiencias de prueba y siguieron siendo el estándar de la compañía por esa razón.
Gallimimus es relativamente colorido, con tonos de piel que van desde el melocotón claro hasta todos los tonos de naranja e incluyen varios bronceados y marrones terrosos. Similar al Velociraptor, el patrón en el lado dorsal es común, con rayas, manchas y patrones conocidos. El color de estos patrones suele ser más oscuro que el del cuerpo principal; púrpura o azulado es común para las rayas, mientras que las manchas tienden a ser de color naranja quemado o marrón, y las alforjas suelen ser de color marrón claro o lavanda descolorida. El patrón generalmente ocurre en los hombros y desaparece hacia las caderas, aunque las rayas en el cuello o la cola no son infrecuentes. Por lo general, hay un sombreado de color beige o crema, y el cuello suele ser más gris, mientras que la cabeza puede ser más amarilla.
")
")
Crecimiento[]
La evidencia fósil muestra que la punta del codo de Gallimimus se vuelve más prominente con la edad, mientras que el hocico se alarga. Como en muchos animales juveniles, los ojos y el cráneo son proporcionalmente más grandes a edades tempranas. Cuando el animal eclosiona, tiene proporciones de patas no muy diferentes a las del adulto adulto, y es capaz de correr a una velocidad decente desde una edad muy temprana. Esta es una adaptación vitalpara un animal cuya principal defensa es dejar atrás a sus depredadores. Los brazos, por el contrario, son más cortos en los jóvenes que en los adultos. En los ejemplares extintos, se han observado directamente los estadios juveniles y subadultos, lo que demuestra los cambios de coloración que ocurren también a lo largo de la vida. Los colores de los juveniles jóvenes son más claros que los de los adultos, generalmente con un color marrón simple o naranja espolvoreado y rayas mínimas.
Si bien los Gallimimus clonados carecen de plumas, la evidencia fósil sugiere que las estructuras de plumas en forma de alas en los brazos habrían crecido cuando el animal alcanzó la madurez. Probablemente se usaron durante las exhibiciones de cortejo.
Los colores cambian drásticamente durante la adolescencia, con manchas horizontales plateadas o blancas que aparecen a los lados del cuerpo mientras que el color general se oscurece. Este color puede ayudar a los jóvenes y sub-adultos mayores a esconderse en ambientes boscosos, similar a los patrones de manchas marrones y blancas en los cervatillos de los ciervos. Las manchas parecen convertirse en rayas a medida que el animal envejece. Una vez que se alcanza la edad adulta, estas vetas blanquecinas se transforman en los colores del animal maduro, lo que probablemente indica madurez sexual. Se desconoce la tasa de crecimiento, pero parece ser bastante rápida y es probable que la edad adulta se alcance en unos pocos años. InGen ha utilizado históricamente fórmulas de hormonas que aceleran el crecimiento para provocar la madurez temprana, lo que hace que sea aún más difícil determinar exactamente qué tan rápido crecería naturalmente un Gallimimus.
Dimorfismo sexual[]
Hasta el momento no existe una descripción concreta del dimorfismo sexual en Gallimimus, pero se ha sugerido que los patrones de color pueden diferir entre machos y hembras. En general, los dinosaurios machos tienen colores más oscuros y vivos que las hembras, lo que sugiere que los colores naranja quemado y las manchas oscuras pueden pertenecer con mayor frecuencia al macho Gallimimus, mientras que los colores marrón melocotón y tierra pertenecen a las hembras.
Comportamiento[]
Patrones de actividad[]
Este terópodo es diurno; no parece estar activo durante la noche y probablemente busca lugares protegidos para dormir. A pesar de sus grandes ojos, no parece estar adaptado a la oscuridad. Se ve con mayor frecuencia durante las horas de la mañana, y parece disminuir alrededor del mediodía. Muchos dinosaurios descansan durante el calor del día y se consideran catérmicos; esto no está confirmado específicamente en Gallimimus. No se ha visto a menudo por la noche y permanece inactivo por la noche a menos que se le moleste.
Dieta y comportamiento alimenticio[]
Aunque con frecuencia se clasifica entre los herbívoros, el Gallimimus es un omnívoro y se alimenta tanto de animales como de plantas. Su anatomía significa que está lejos de ser un cazador de caza mayor, y rara vez mata a algo que pueda resistir mucho. Probablemente se alimenta principalmente de insectos, gusanos y pequeños reptiles como carne. En lo que respecta a las plantas, prefiere la vegetación blanda, como hojas y frutos, en lugar de material leñoso resistente. Se sabe que se alimenta de plantas ricas en lisina, como la soja, aunque es probable que se alimente de forma oportunista y coma todo lo que pueda encontrar en su entorno. Algunos científicos creen que podría ser capaz de incursionar como patos para alimentarse de pequeños invertebrados acuáticos, o que podría comerse los huevos de otros animales.
La parte frontal de su cuello es flexible, lo que permite un buen rango de movilidad para que Gallimimus pueda alcanzar las fuentes de alimento tanto en posiciones altas como bajas. Esto también le ayuda a llegar a más áreas cercanas a sí mismo sin tener que moverse tanto. No tiene dientes y debe tragar la comida entera, pero tiene laminillas similares a las de las aves acuáticas modernas. Estos se asemejan a dientes y pueden usarse para ayudar a este dinosaurio a cortar trozos de hojas y frutas, así como a agarrar presas de pequeños animales que luchan. Algunos paleontólogos también han sugerido que, al igual que ciertas aves acuáticas, las laminillas pueden usarse para meterse en el agua a nivel de la superficie y tamizar pequeños crustáceos y otros invertebrados. Sin embargo, esto no sería suficiente para sostener a un animal de este tamaño solo. Algunos paleontólogos han estimado que, dependiendo de su metabolismo, un Gallimimus podría necesitar comer entre 0,15 y 7,36 libras (0,07 y 3,34 kilogramos) de comida por día.
Los fósiles de ornitomimidos han mostrado la presencia de gastrolitos, pequeñas piedras ingeridas por animales herbívoros que no pueden masticar para triturar plantas en el tracto digestivo. Si bien esto no se ha visto directamente, el sitio web de InGen IntraNet describe la geofagia en Gallimimus cuando el dinosaurio usa sus garras para rasgar la capa superior del suelo y exponer las fuentes de alimento. Mientras come lo que ha descubierto o arrancado del suelo, también se traga tierra y pequeñas rocas. El sitio web sugiere que esto no es intencional, pero si Gallimimus usa gastrolitos como sugieren los fósiles, puede estar tragando piedras a propósito para ayudar en su digestión.
Comportamiento social[]
Gallimimus es un animal de manada, que vive en grandes grupos con fines de supervivencia. Su velocidad y cautela son sus principales defensas contra los depredadores, pero al vivir en grupos más grandes, cada animal individual tiene más posibilidades de no ser presa. Si un Gallimimus detecta peligro, huirá y su pánico se esparcirá rápidamente por todo el rebaño y provocará una estampida. Las bandadas de Gallimimus realizan movimientos evasivos coordinados y cambios de dirección uniformes mientras se desplazan en estampida, lo que hace que sus esfuerzos por huir de los depredadores sean un asunto organizado en lugar de caótico.
No parece haber ninguna jerarquía social diferenciada entre las bandadas de Gallimimus; en lugar de un líder o animal alfa presente, todo el rebaño actúa como una sola unidad. Este es un rasgo común en los terópodos más pequeños, particularmente aquellos que son herbívoros u omnívoros. Muchas aves en bandadas modernas exhiben una estructura social similar, al igual que los pequeños terópodos no aviares como el Compsognathus. Esta falta de un líder individual significa que la única forma de lograr el poder social dentro del rebaño es ser lo suficientemente rápido y ágil para sobrevivir: los lentos y torpes son los primeros en ser asesinados por depredadores en una forma brutal de selección natural.
Esto no quiere decir que los Gallimimus no tengan vínculos verdaderos entre sí. Mientras que los animales más rápidos tienen más probabilidades de sobrevivir, los adultos mantendrán a los juveniles en medio de la bandada durante las estampidas para protegerlos. También hay alguna evidencia de que diferentes adultos pueden tener vínculos emocionales. Durante el incidente de 1993 en Isla Nublar, una bandada de Gallimimus fue perseguida por la Tyrannosaurus del Parque Jurásico, que preparó una emboscada y mató a uno de los animales que tropezaron durante el ataque. Mientras los otros huían, uno de los Gallimimus hizo una pausa y se dio la vuelta, viendo como mataban y comían al otro. El resto de la bandada disminuyó la velocidad, pero no se detuvo ni retrocedió. Si bien el comportamiento solo se observó brevemente, parece que Gallimimus puede formar amistades y angustiarse cuando el daño llega a otros miembros de sus rebaños con los que están vinculados.
Gallimimus también es conocido por ser juguetón. Su juego favorito, naturalmente, son las carreras. Cuando todo esté seguro en su entorno, cargarán a través de campo abierto en pequeños grupos y mostrarán su velocidad. Presumir de esta manera no solo demuestra sus habilidades de supervivencia, sino que también anima a otros Gallimimus a mejorar su fuerza y velocidad al correr. Esta competencia conduce a un ejercicio frecuente que asegura que la bandada esté mejor preparada colectivamente para sobrevivir a un ataque y les da a estos dinosaurios una forma de estimulación. Se sabe que compiten con vehículos de la misma manera, y probablemente también disfruten compitiendo contra otras especies de animales.
Reproducción[]
Actualmente no se sabe mucho sobre la reproducción de Gallimimus, aparte de que ponen huevos y se aparean mediante la cloaca como la mayoría de los dinosaurios. Los huevos de dinosaurios de tamaño mediano, como este, generalmente se incuban durante un período de unos meses, y la mayoría de las especies de dinosaurios practican el cuidado parental extendido. Los terópodos suelen tener huevos ovoides, una adaptación para que sea más fácil mantenerlos en un lugar sin rodar. En la prehistoria, los Gallimimus tenían plumas y es posible que pudieran criar sus huevos, pero los especímenes de InGen no tienen plumas y probablemente sean menos capaces de empollar.
Los gallimimus más jóvenes muestran una coloración claramente diferente a la de los adultos, lo que sugiere que el color juega un papel social. Las crías y los ejemplares más jóvenes tienen un color simple y liso, mientras que los adolescentes tienen rayas laterales blanquecinas. Los adultos pueden tener rayas verticales y manchas, y tienden a tener un color base más variable que los juveniles también. Todos los adultos protegerán colectivamente a los juveniles y adolescentes manteniéndolos en el medio de la bandada durante las estampidas, lo que hace menos probable que los depredadores se aprovechen de ellos. La mortalidad infantil parece ser alta ya que los juveniles rara vez se ven en la naturaleza, por lo que mantener seguros a los que sobreviven es una alta prioridad.
No está claro a qué edad se alcanza la madurez o cuándo es la temporada de reproducción. Los Gallimimus maduros probablemente se anuncian a posibles parejas utilizando su coloración y su fuerza física. Competir unos contra otros es un comportamiento de juego común en esta especie, y dado que esto anuncia su velocidad y agilidad, sería un comportamiento de cortejo adecuado. En Gallimimus, ser rápido significa mantenerse con vida, por lo que este es un rasgo deseable en una pareja.
Comunicación[]
En su mayor parte, Gallimimus es una especie tranquila. Al estar más abajo en la cadena alimentaria y vivir a menudo en ambientes con abundantes depredadores, no hace mucho ruido. Probablemente se comunica principalmente con el lenguaje corporal, ya que su cuello largo y sus poderosas piernas pueden usarse para realizar una serie de movimientos distintivos. Por ejemplo, se cree que inclinar la cabeza hacia otra criatura mientras corre es un desafío para correr.
Se conocen algunas vocalizaciones, la mayoría de las cuales se utilizan al correr. Al acercarse a obstáculos (o animales más lentos), Gallimimus emitirá chillidos y graznidos, que no hacen cuando mantienen el rumbo. Esto significa que estos ruidos probablemente se utilizan para advertir a los otros Gallimimus antes de hacer una corrección de rumbo, lo que ayudaría a evitar colisiones durante las estampidas. Chocar o tropezar sería una consecuencia fatal para este ágil pero frágil dinosaurio, por lo que debe evitarse a toda costa.
Ecología[]
Siendo un omnívoro con preferencia por las hojas y las frutas, este dinosaurio puede ser útil para algunas plantas y perjudicial para otras, e incluso puede ayudar y dañar simultáneamente. Comer las hojas de las plantas es un comportamiento depredador, pero cuando se alimenta de frutos, puede esparcir semillas a nuevos entornos. Por tanto, a largo plazo, Gallimimus puede alterar el tipo de vida vegetal en su hábitat.
Es relativamente pequeño, en comparación con muchos otros dinosaurios, por lo que no puede causar mucho daño a plantas más grandes como los árboles. Gallimimus, por ejemplo, se ha visto en ambientes de secuoyas; poco puede hacer para afectar a un árbol tan grande cuando está completamente desarrollado. La flora más pequeña, como helechos y arbustos, puede ser pisoteada o destrozada por este dinosaurio. Es probable que se vean pastos en su hábitat en lugares donde comúnmente se alimenta y deambula. Incluso puede destrozar la vida vegetal sin darse cuenta mientras se alimenta de pequeños animales como los insectos.
Gallimimus es una presa común para numerosos carnívoros. Los depredadores confirmados incluyen al Tyrannosaurus y al Velociraptor; mientras que Velociraptor es lo suficientemente rápido y ágil como para matar fácilmente a un Gallimimus, el Tyrannosaurus carece de la resistencia suficiente para perseguir a estas criaturas. En cambio, es común que el T. rex cace utilizando emboscadas, asustando a las bandadas de Gallimimus para que se muevan y luego sorprendiéndolos desde un escondite. Fue una de las víctimas del Scorpius rex en 2016, con una perdida confirmada. Otros depredadores potenciales incluyen Allosaurus, Teratophoneus, Carnotaurus y Pteranodon; en particular, Pteranodon probablemente se alimenta de gallimimus juveniles, y el Carnotauro, de rápidos movimientos, probablemente podría capturar a un gallimimus. Otros carnívoros de las mismas áreas incluyen al Dimorphodon, que podría amenazar a un juvenil pero probablemente no a un adulto, y Baryonyx, que probablemente sería un depredador de emboscada. El diminuto Compsognathus es menos peligroso, aunque sus mordeduras levemente venenosas podrían ralentizar a un Gallimimus lo suficiente como para hacerlo vulnerable a amenazas mayores. Grupos más grandes de Compsognathus probablemente podrían matar a un joven o un adolescente débil.
Tiene pocas defensas importantes contra los depredadores. Aunque sus patas son fuertes y podrían dar una patada sólida, esto requeriría que se acercara bastante a un depredador; en lugar de luchar, huirá a gran velocidad. Es más probable que los depredadores eliminen a los miembros más lentos y débiles de la bandada, por lo que cuanto mayor sea la población de Gallimimus, mayor será la probabilidad de supervivencia de cada individuo. Un Gallimimus solitario puede intentar intimidar a un depredador con un chillido o un silbido, pero esto no suele ser muy eficaz y el animal debe recurrir a la huida a gran velocidad.
Otra forma en que puede permanecer más seguro es vivir entre especies de presa más grandes y resistentes. A menudo se le ve cerca de saurópodos, como Mamenchisaurus, Brachiosaurus y Apatosaurus. Estos grandes herbívoros son a menudo los primeros en detectar un depredador que se acerca y están bien defendidos contra todos los carnívoros, excepto los más grandes. También se les ha visto convivir con especies comoOuranosaurus, Pachycephalosaurus y Stygimoloch, que son territoriales y capaces de defenderse, y el voluminoso hadrosaurido Parasaurolophus (que presenta un objetivo más grande y lento, más tentador para un depredador hambriento). Se sabe que Edmontosaurus, otro hadrosaurido, vivió en hábitats compartidos con Gallimimus. Los dinosaurios blindados también son buenos para convivir por sus capacidades defensivas; Ankylosaurus, Peloroplites, Stegosaurus, Sinoceratops y Triceratops son todos conocidos de territorios que también incluyen Gallimimus. De todos estos, Triceratops es probablemente el que representa un mayor riesgo, ya que es el más territorial y acosará a otros dinosaurios para proteger sus fuentes de alimento. Los dinosaurios juveniles de otras especies han muerto de hambre a causa del agresivo Triceratops y el apacible Gallimimus. tiene la desventaja añadida de ser un terópodo.
Se desconoce si este dinosaurio come huevos de otras especies, como han sugerido algunos paleontólogos; si lo hace, a menudo vive junto a muchos otros animales a los que podría robar de manera confiable. Su velocidad lo ayudaría aquí, ya que tendría que ser evasivo para evitar ser pisoteado, corneado o mordido por un padre protector.
Gallimimus también está sujeto a parásitos y enfermedades, como todos los animales. En la era Mesozoica, fue picado por mosquitos , que es como InGen obtuvo su ADN. Si bien no está confirmado, es posible que los mosquitos modernos lo piquen hoy. Según Jurassic World: Evolution, es particularmente susceptible a la influenza aviar, que generalmente es causada por el virus de la influenza A. Como especie de presa común, Gallimimus sería un huésped intermedio ideal para parásitos y patógenos, especialmente porque cualquier infección que lo ralentice sería rápidamente fatal.
Hábitat[]
En la prehistoria, este dinosaurio vivía en llanuras aluviales semiáridas con lluvias estacionales. La evidencia fósil sugiere que vivía cerca de ríos y otros cuerpos de agua dulce. Desde que fue recreado, se ha adaptado bien a una variedad de ambientes, principalmente áreas cálidas como pastizales tropicales, regiones semiáridas, bosques nubosos y bosques de coníferas.
Isla Sorna[]
Los primeros Gallimimus clonados fueron criados originalmente por InGen en Isla Sorna, probablemente en algún momento a principios de la década de 1990. Esto se estima porque los guiones anteriores de Jurassic Park implican que poblaciones de Gallimimus exclusivamente femeninas se estaban reproduciendo, lo que significa que eran capaces de efectuar la protoginia (esto, sin embargo, se cambió a Velociraptor en la película final, lo que indica que puede no ser canon). Dado que los desarrollos recientes revelaron que los genes que codifican la protoginia se integraron por primera vez accidentalmente en los genomas de los dinosaurios en 1991, Gallimimus tendría que haber sido creado después de esto.
En cualquier caso, este terópodo probablemente fue creado dentro del Complejo de Embriología, en la parte occidental de la isla. Aquellos que fueron aprobados para la exhibición en el parque, fueron enviados a Isla Nublar una vez que crecieron lo suficiente, mientras que otros se mantuvieron en el Sitio B para su investigación.
Después del incidente de 1993 en Isla Nublar, la producción en Isla Sorna cesó debido a la creciente crisis financiera de InGen. En 1995, el huracán Clarissa provocó una evacuación completa, y los animales se soltaron a la naturaleza bajo el supuesto de que morirían debido a la contingencia de lisina. Al igual que los otros dinosaurios, Gallimimus sobrevivió alimentándose de fuentes alimenticias ricas en lisina, que existen en abundancia en la naturaleza. En 1993, InGen registró veinte Gallimimus viviendo en Isla Sorna.
No se sabe mucho sobre la población entre 1993 y 1997, aunque las imágenes térmicas satelitales de la isla revelaron una población de Gallimimus en el este a principios de 1997. Durante las expediciones de 1997 a Isla Sorna, se vieron entre quince y treinta adultos en un sendero de caza del noreste; dos de ellos fueron capturados por InGen y retenidos en un campamento en el norte. Durante la primera noche del incidente, los activistas liberaron a los animales capturados, que huyeron al bosque circundante.
Nuevos animales fueron introducidos ilegalmente a Isla Sorna entre finales de 1998 y mediados de 1999, alterando el ecosistema, ya de por sí afectado, de la pequeña isla. Aunque el Gallimimus es capaz de comer una amplia variedad de alimentos, la competencia que ahora enfrentaba era intensa. Dejado a sus propios dispositivos, puede haber sobrevivido a algunos de sus competidores, pero el interés humano intervino de todos modos; Entre enero de 2004 y mayo de 2005, se iniciaron operaciones dentro de InGen para reubicar a los dinosaurios restantes de Isla Sorna a Isla Nublar. Supuestamente, esto se concluyó cuando Jurassic World abrió sus puertas el 30 de mayo de 2005 y no quedan dinosaurios en Isla Sorna. Sin embargo, la continuación de la actividad de caza furtiva cerca del archipiélago de Las Cinco Muertes sugiere que algunos especímenes de interés todavía pueden vivir allí.
Isla Nublar[]
Este dinosaurio se introdujo por primera vez en Isla Nublar a principios de la década de 1990, para formar parte de la fauna de Jurassic Park; los guiones anteriores implican que se encontraba entre las creaciones más recientes en 1993. Se le asignó un prado más pequeño y estrecho en las praderas de la isla, centradas en el Río de la selva. Prácticamente toda su frontera norte se puso en contacto con lo que originalmente estaba destinado a ser el recinto del Velociraptor (después de que las rapaces se consideraran inadecuadas para la exhibición en el parque, este recinto estubo destinado a contener al más manejable Herrerasaurus en su lugar). Solo una pequeña parte de la frontera noroeste entró en contacto con el prado principal de herbívoros. El resto de su hábitat bordeaba la carretera principal de turismo, separada de ella por un foso de hormigón ; esta era la única parte del prado que no estaba acordonada por una cerca eléctrica de 24 pies. Algunos mapas implican que este dinosaurio se habría exhibido eventualmente en la región sur de la isla.
Para el 11 de junio de 1993, había al menos 36 Gallimimus en Isla Nublar; seis de ellos eran subadultos. Los guiones anteriores de la película implican que estos dinosaurios podrían reproducirse por protoginia, lo que implica la existencia de machos terminales; sin embargo, esto nunca ha sido confirmado en el canon. El 11 de junio, se cortó el suministro eléctrico a los sistemas de seguridad de la isla durante un caso de espionaje corporativo, y durante el apagón subsiguiente, la bandada de Gallimimus escapó de su recinto. Parece que se dirigieron al prado de herbívoros contiguo, habitado por Parasaurolophus y Brachiosaurus. Sin embargo, no fueron los únicos terópodos que se mudaron a este prado; fueron seguidos por la Tyrannosaurus de la isla, que atacó y mató a un Gallimimus adulto durante la mañana del 12 de junio. Ese día se restauró la energía de las cercas eléctricas, pero el daño a la infraestructura del parque ya era lo suficientemente grande como para permitir que los dinosaurios deambularan por donde quisieran.
Cuando InGen dejó de monitorear la isla en 1993, la población de Gallimimus se había reducido a 24. La depredación del Tyrannosaurus continuó reduciendo su número; para el 5 de octubre de 1994, solo quedaban nueve ejemplares.
Se desconoce cuántos Gallimimus, si es que había alguno, todavía estaban vivos en abril de 2002, cuando InGen se instaló en Isla Nublar nuevamente bajo el ala de Masrani Global Corporation. Los restos óseos de un adulto fueron encontrados en el área del valle central, donde había muerto el primer animal asesinado en 1993, pero no se sabe si se trataba del mismo individuo. Si alguno todavía estuviera vivo, Masrani Global e InGen los habrían protegido de una mayor depredación, y eventualmente los habrían trasladado al valle central en 2004, cuando el parque fue reconstruido como Jurassic World. Este valle se incorporó gradualmente a la atracción de la girosfera y se designó una zona específica como el Valle de Gallimimus. Para 2014, la población de Isla Sorna fue enviada a Isla Nublar y se inició la cría de este animal en el Laboratorio de Creación Hammond.
La población volvió a aumentar; en agosto de 2004 había al menos tres, y probablemente más, ejemplares, pero para diciembre de 2015, había al menos 48 adultos en el valle. Los jóvenes se mantuvieron en el zoológico de mascotas Gentle Giants hasta que tuvieran la edad suficiente para vivir solos; en diciembre de 2015 había al menos 17 menores, todos de edad bastante temprana.
El 22 de diciembre de 2015, Jurassic World experimentó un grave incidente de seguridad que cerró permanentemente el parque. Los Gallimimus ahora estaban solos y, una vez más, estaban sujetos a depredadores. Se desconoce si los jóvenes fueron liberados al valle por sus cuidadores, o si alguno sobrevivió; ninguno se vio en los años siguientes. Sin embargo, con una población inicial de más de 48 adultos y muchas más especies de presas para que los depredadores cazaran, el Gallimimus tenía más posibilidades de sobrevivir.
El valle central, aunque era un hábitat privilegiado para esta especie amante de los pastizales, ya no era seguro. Se vieron obligados a buscar un hogar en otro lugar, muchos de ellos emigraron al norte. La mayoría parece haber habitado la región noroeste de Isla Nublar, en los bosques cercanos al Monte Sibo. Al menos uno fue asesinado en junio de 2016 por un Scorpius rex, que diezmó las poblaciones de numerosas especies en la isla.
Este hábitat no permanecería tranquilo por mucho tiempo; a principios de 2017, la actividad tectónica volvió a despertar el volcán, conectándolo con cámaras de magma de alta presión debajo de la Isla Nublar. La presión se acumuló debajo de la isla durante el año siguiente, y esto tuvo efectos devastadores en el ecosistema. Vivir tan cerca del volcán puso a Gallimimus en peligro, lo que puso aún más estrés a la población. Los restos óseos de un subadulto fueron vistos cerca del Radio Bunker 02-17 el 23 de junio de 2018, aunque parecía bastante intacto y probablemente no murió en un ataque. En ese momento, se vieron 34 adultos cerca del lado este del volcán, y otros seis adultos fueron capturados por mercenarios liderados por el cazador Ken Wheatley antes de ese día. Esto da una población total de al menos 40 a partir de ese momento, una disminución de los 48 adultos y 17 jóvenes que vivían en el parque cuando cerró.
El 23 de junio, el Monte Sibo hizo erupción y devastó Isla Nublar. Dado que los Gallimimus estaban tan cerca del volcán, fueron inmediatamente afectados por su erupción; los gases tóxicos, las nubes de cenizas y las bombas volcánicas mataron a muchos de ellos. Algunos saltaron por los acantilados del noroeste y cayeron cientos de metros al mar. Muchos probablemente se ahogaron o murieron en el impacto al caer desde tal altura. Aquellos Gallimimus que sobrevivieron se habrían enfrentado al último desafío en la supervivencia, con la vida vegetal de la isla en su mayoría incinerada y las pequeñas presas muertas de hambre, asesinadas o envenenadas. Si bien algunos carroñeros oportunistas aún pueden sobrevivir en Isla Nublar permaneciendo en el sur, viviendo de plantas resistentes y detritos oceánicos, el tamaño corporal más grande de Gallimimus hace que sus perspectivas sean sombrías. Es poco probable que quede alguno en la isla en la actualidad, ya que la actividad volcánica aún continúa; se cree que los únicos supervivientes son los que fueron trasladados al SS Arcadia.
El último Gallimimus sacado de la isla se cargó en el Arcadia durante la erupción, se registró en el manifiesto a las 13:43 y fue cofirmado por Moti Biran. Este animal se mantuvo en el Contenedor # 25-1009-5810 (Carga # 84112). Pesaba 190 kilogramos, lo que lo hacía notablemente bajo de peso.
En el mundo actual[]
Había planes para exhibir Gallimimus en Jurassic Park: San Diego, que se habría inaugurado en 1997, pero esto nunca se completó debido a las luchas internas entre John Hammond y Peter Ludlow.
Actualmente no hay evidencia de que Gallimimus haya sido traído al continente antes de 2018, aunque se sabe que la caza furtiva ha ocurrido tanto en el Archipiélago de Muertes como en la Isla Nublar. El primer caso confirmado de remoción de este dinosaurio de las islas, donde fue creado, es el 24 de junio de 2018; al menos seis adultos fueron sacados ilegalmente de Isla Nublar por un equipo mercenario liderado por Ken Wheatley y contratado por Eli Mills, de la Fundación Lockwood. Estos animales fueron capturados y transportados a través de SS Arcadia a la Residencia Lockwood, cerca de Orick, California, para ser vendidos en el mercado negro.; sin embargo, no se ha confirmado que se vendiera ninguno esa noche. Durante el incidente en la mansión, una fuga de cianuro de hidrógeno puso en peligro a los dinosaurios capturados pero no vendidos, y para evitar que se extinguieran por completo, Maisie Lockwood los soltó en el bosque circundante. La pequeña bandada de Gallimimus superviviente ahora vive en el oeste de América del Norte, y se desconoce su ubicación actual.
Canon Cinematográfico[]
Películas[]
Jurassic Park[]
En el segundo día del Incidente de Isla Nublar, el Dr. Alan Grant, junto con Lex y Tim Murphy presenciaron una bandada de Gallimimus en su recinto huyendo de la Tyrannosaurus rex de Jurassic Park. El T. rex logró capturar y matar a un desafortunado individuo, al que emboscó cuando el rebaño corría por seguridad.
Durante la segunda película, se pueden apreciar varios ejemplares en un sendero de casa; dos adultos Gallimimus fueron capturados por los cazadores de InGen para el Parque Jurásico que Peter Ludlow pretendía crear en San Diego, pero fueron liberados la noche siguiente por Nick Van Owen y la Dra. Sarah Harding, junto con el resto de los dinosaurios cautivos de los cazadores.
Gallimimus hace varios cameos a lo largo de la historia; se ve a una manada de estos terópodos en el valle de Gallimimus, que pasan junto a un vehículo de zafari. También se ven a algunos de los menores que vivían en el zoológico de mascotas Gentle Giants.
Jurassic World: Fallen Kingdom
Fueron vistos corriendo junto a muchas otras especies lejos del Monte Sibo. Algunos Gallimimus también fueron capturados y llevados a la Mansión Lockwood para una subasta, pero gracias a Claire Dearing y Maisie Lockwood, eventualmente escaparían de la residencia junto con los otros dinosaurios.
Series[]
Jurassic World: Camp Cretaceus
Varios ejemplares de Gallimimus se ven durante la tercera temporada de la serie. En particular, Darius Bowman sigue a una manada para documentar la especie en su diario de campo, interactuando con un par de ejemplares. También descubre un ejemplar parsialmente consumido que cae de lo alto de un árbol, lo que, aunque de forma indirecta, le revela la existencia del Scorpius rex.
Libros[]
Gallimimus aparece como una de las cinco especies que residen en el Valle de la Girosfera. Solo se los ve una vez en la historia de forma directa, cuando tres de ellos corren junto a un jeep.
Canon Literario[]
The Lost World
En El mundo perdido, se hace una referencia indirecta a Gallimimus, lo que demuestra que existe en Isla Sorna en el universo de la novela. Su nombre (Gallimimus bullatus) está impreso en un documento dirigido al Dr. Henry Wu, entre las docenas esparcidas por todo el complejo abandonado de InGen.
Videojuegos, comics y otors multimedios[]
Gallimimus aparece en las lineas de juguetes con un único modelo y una cría para uno de los humanos. Aparece en el comic de Topps, y en los Juegos Jurassic Park Rampage Edition donde Alan Grant puede montarlo para el nivel de la sahavana, Jurassic Park arcade donde aparecen en estampida, The Lost World Jurassic Park Arcade haciendo cameo en el nivel de la sahavana, Dinosaur Battles como el dinosaurio mas debil del juego, Jurassic Park Operation Genesis donde podemos criar a este animal, en el juego es retratado como un herbivoro pequeño y debil que sirve de alimento a todos los carnivoros, su popularidad es de 2 estrellas, Jurassic Park 3 Park Builder como un carnivoro pequeño de bajo nivel, Jurassic Park DVD Explorer, Jurassic Park The Game haciendo un cameo leve, Jurassic Park DNA Factor y Jurassic Park Island Attack. Es nombrado entre las especies que viven en Isla Sorna durante los eventos de Jurassic Park 3, inclusive hay artes conceptuales promocionales de la pelicula que muestran a esta especie.
Aparece en el folleto de especies de Jurassic World asi como en el trailer en donde se lo ve en estampida junto a un bus de safari, tiene su propia atraccion "Gallimimus Valley" y pueden ser vistos sus bebes en el Zoo de Gigantes Gentiles. Ademas esta presente en el plantel del Jurassic World The Game para IOS donde aparece con dientes y sus sonidos son semejantes a los Velociraptor lo que hace la reconstruccion bastante inexacta, ademas puede formar un hibrido el Ophiacomimus (Ophiacodon + Gallimimus). Un diseño sin plumas pero que corrige las muñecas pronadas en la anatomia de Gallimimus fue creado para el juego cancelado de Jurassic World. Tambien forma parte de los dinosaurios en el juego LEGO Jurassic World. La linea de juguetes de Hasbro Jurassic World Hybrids presenta al hibrido Pteramimus (Pteranodon + Gallimimus). El juego Lego Jurassic World tiene su modelo de Gallimimu. Regresa para Jurassic World Fallen Kingdom, Mattel ya saco una figura de accion de este dinosaurio en varias variantes y tambien es parte del plantel de dinosaurios para el juego Jurassic World Evolution donde es el herbivoro tipico que sirve de alimento a los carnivoros de la isla y no posee defensas mas que correr, aunque de por si la animacion del juego le quita la opcion de escapar, una vez que la secuencia inicia ya el destino del animal esta sellado y tambien aparece en el Jurassic World Facts de Mattel, y Jurassic World: Alive para dispositivos moviles donde ademas forma al hibrido Monomimus (Monolophosaurus + Gallimimus) que a su vez si une su genoma con el del Darwinopterus forman al hibrido Pterovexus, tambien puede unir su genoma con la Aguila de Haast del tipo GEN 2 y formar al Aquilamimus (Harpagornis moorei GEN 2 + Gallimimus).
Su aparicion en la pelicula Jurassic Wold Fallen Kingdom se reduce a cameos, en las escenas de la estampida de la erupcion volcanica del monte Sibo de Isla Nublar donde en el caos se pueden ver varios ejemplares dispersos escapando y pasando cerca de Franklin Webb, Zia Rodriguez, Claire Dearing y Owen Grady, se ve a un ejemplar luchar con los cazadores que lo someten y lo encarcelan y en el escape de los dinosaurios de la mansion Lockwood, al igual que los demas dinosaurios escapa hacia el bosque con paradero desconocido.
En 2020 se creo el DLC oficial de Minecraft Jurassic World donde ademas de aparecer este dinosaurio puden crearse hibridos como Symmachosophus (Parasaurolophus + Velociraptor + Stygimoloch + Gallimimus).
Galería[]
")
")
")
")
")
.jpeg "WhatsApp Image 2018-04-06 at 14.53.36 (1).jpeg (108 kB)")
.png "Descarga (5).png (83 kB)")
")
.jpeg "WhatsApp Image 2018-04-20 at 06.36.37 (1).jpeg (108 kB)")
.jpeg "WhatsApp Image 2018-04-23 at 20.53.56 (1).jpeg (106 kB)")
")
")
")
")
.jpg "Galli s-l300 (2).jpg (9 kB)")
")
")
")
")
")
")
")
")
")
")
")
")
")
")
")
")
")
")
.png "Gallimimus (5).png (64 kB)")
")
")
")
")
")
")
")
")
")
")
")
.jpg "Galli (2).jpg (117 kB)")
")
")
")
")
")
")
")
")
")
")
")
")
")
")
")
")
")
")
")
")
")
")
")
")
")
")
")
")
")
")
")
")
")
")
")
")
")
")
")
.jpeg "WhatsApp Image 2020-11-21 at 21.22.08 (1).jpeg (189 kB)")
")
")
")
")
")