{kind=link}
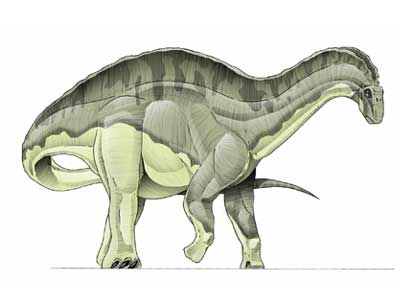
Dicraeosaurus ("reptil bifurcado") es un género con dos especies conocidas de dinosaurios saurópodos dicreosáuridos, que vivieron a finales del período Jurásico, hace aproximadamente hace 156 a 145 millones de años, entre el Oxfordiense y el Titoniense, en lo que hoy es África. Su nombre se debe al notable desarrollo de las espinas de las vértebras de la espalda y cuello. Fue descrito por primera vez por el paleontólogo alemán Werner Janensch en 1914, en su monografía sobre la fauna de la formación de Tendaguru. En esta nombró las dos especies conocidas, Dicraeosaurus hansemanni y Dicraeosaurus sattleri.
Descripción[]
Dicraeosaurus medía entre 13 y 20 metros de largo y hasta 6 de alto, con un peso de 15 toneladas. Poseían una larga cola estrecha, un dorso con espinas, patas macizas y una cabeza larga con largos incisivos en sus mandíbulas, el cuello estaba construido con solo 12 vértebras, siendo más corto que los de los otros saurópodos de su época. A diferencia de la mayoría de los diplodocoides Dicraeosaurus tenía una cabeza relativamente grande puesto que su cuello era más corto y más ancho. También careció la cola en forma de látigo que los diplodócidos tenían. Su tamaño era también diferente. La característica más notable y la que le da el nombre son las espinas dorsales de las vértebras. Estas no eran rectas como en otros miembros de la familia. Cada una tenía una forma de Y, como una bifurcación. Estás espinas sirvieron como puntos de inserción para poderoso músculos del cuello. En la parte superior del hueso del muslo en el lado lateral hay un cuarto trocánter corto.
Dicraeosaurus tenía hasta 20 metros de largo, de los cuales 2,4 metros pertenecen al cuello. Numerosos intentos de estimar su masa arrojaron resultados variados, colocando la masa de D. hansemanni entre 10 y 40 toneladas. En 1988, el cálculo se realizó sobre la base de esqueleto de Berlín, para que parte de los hueso utilizado para reconstruir vino del mismo individuo y los faltantes sustituidos por animales de similar tamaño de la misma especie, así como moldes de yeso. Los cálculos se basaron en estimaciones precisas del área y el volumen de las partes individuales del cuerpo del animal. Su volumen total fue de 12,81 m³, con 9.38 m³, más de 73%, se atribuyó a la torso y sólo 0,05 m³ a la cola, 4 ‰ del total, la cola misma ocupaba más de un metro cúbico, y el cuello ¾ m³. Se obtuvieron resultados similares para la superficie del cuerpo, el valor total de 46,4 m² casi 56% era tronco, 11,7 % la cola, y 9,4 % las patas traseras, solamente 2,8 % las delanteras y un poco más de 7 % el cuello. La pequeña cabeza era de 9 ‰. Estos cálculos permitieron estimar la masa de un dinosaurio en 12,8 toneladas. Cabe señalar que la masa aproximada con el mismo método para Giraffatitan brancai, reconocido en el momento de la medición como una especie de Brachiosaurus ascendió a más de 74 toneladas. Los cálculos ya mencionados también han sido usados para estimar la tasa metabólica de ambos animales. El representante Dicraeosaurus consumió 400 MJ por día, mucho menos que Giraffatitan con pulmones seis veces más grandes. En Dicraeosaurus contenían 974 litros de aire. Los cálculos muestran que 641 litros de sangre se bombeaban por un corazónque pesaba 48 kilogramos latiendo 23 veces por minuto. Dicraeosaurus creció rápidamente y sin interrupción hasta su tamaño máximo. Alcanzando el 80 % de este tamaño máximo, maduraba sexualmente. Este alto valor, en comparación con otros saurópodos, indica que el animal habría dejado de crecer antes que sus parientes. Esta hipótesistambién se sostiene por la pobre neumatización de sus huesos.
Sus dientes con forma de clavijas son ideales para alimentarse de las plantas. Tanto Dicraeosaurus, como Giraffatitan muestran signos de cizallamiento especialmente en su parte superior y en menor medida en el medio y atrás. Al igual que la estructura del cráneo, que apuntan a la inclinación de los movimientos de las mordidas durante la comida,10 otros autores ver aquí los movimientos de la parte delantera-trasera,11 que se producía en diplodocoides más adaptado para cortar alimentos que para arrancarlos como anteriores saurópodos. No se puede descartar que, al igual que Diplodocus, perteneciente a la misma superfamilia, el dinosaurio descrito desarrollara dientes incisivos doblados hacia adelante y mandíbulas frontales.
Entre los huesos frontales y parietales, y entre los huesos parietales y supraorbitales, hay fontanelas cuyas dimensiones están en la proporción de 3:2. Esto último no se encuentra, excepto en Dicraeosaurus y Amargasaurus. Ambos tipos se caracterizan por un apéndice alado muy largo, hacia adelante y ligeramente ventral. El hueso supraorbital es estrecho, está dispuesto verticalmente. En la parte posterior del cráneo hay una bóveda normal, a diferencia de otros sauropodomorfos. Hay un peine en la parte posterior de la cabeza.
Las vértebras cervicales de este animal, que en Dicraeosauridae no es superior en el número a 12, mostraron numerosos espacios aéreos,12 más desarrollado en la parte posterior del cuello. Algunos de ellos incluso tenían una profundidad significativa y rodeaban la parte central del cuerpo vertebral sobre el que se apoyaba. Su disposición se asemeja a la vista en Brachiosaurus. Sin embargo, no se encontraron cámaras de aire internas, que tampoco están presentes en otros huesos de Dicraeosaurus. Como resultado, el peso del cuello del animal disminuyó en un 6Hipótesis (método científico)%. Se suponía que formaban sacos aéreos, estructuras que también se encuentran en las aves. La columna vertebral estaba arqueada, pero no había doblez entre el cuello y el torso. La superficie cóncava sobre la vértebra cervical ventral junto a las costillas del espacio cervical podría acomodar las arterias carótidas. Las diferencias en el peso corporal de la cabeza y cuello confirmaron que en contraste con Brachiosaurus, Dicraeosaurus sostiene el cuello horizontalmente compensando con la cola y el gran tamaño de las extremidades posteriores, en el que la tibia mide los 79 centímetros. Según Martin y colaboradores, el papel decisivo en el mantenimiento del cuello sería un ligamento cervical. Teniendo en cuenta la longitud de los apéndices del cuello, que se piensa que pueden demuestra que los músculos dorsales de Dicraeosaurus fueron mucho mayores que en los diplodócidos.7 Se notó que los animales de hoy que caminan sobre patas estiradas generalmente mantienen el cuello erguido y doblan la cabeza. Esto también debería aplicarse a Dicraeosaurus. Otros expertos creen que el cuello también podría inclinarse, pero no excluyen una posición horizontal. Ambas publicaciones coinciden en que no hay indicaciones de que el cuello esté hacia arriba. De acuerdo con algunos paleontólogos los dinosaurios de cuello largo podían alimentarse de las plantas que crecen en diferentes niveles Según otros expertos, Dicraeosaurus sin embargo, pastaba principalmente en la vegetación baja, y el cuello les permitió llegar lejos a los lado. Se estimó que a pesar de su un mayor cuello Diplodocus o Apatosaurus comida el follaje que se encuentra a una altura de no más de 4-6 metros, el rango de Dicraeosaurus se limitaba más.11 El cuello largo de los saurópodos también podría ser el resultado de la selección sexual.
Las vértebras dorsales delanteras eran más altas y más cortas. Las vértebras caudales proximales se caracterizaron por una pequeña concavidad en el frente y una convexidad en la parte posterior. Generalmente, sin embargo, las vértebras caudales se definen como alargadas, biconvexas. Del mismo modo, se construia la columna vertebral de la cola de los Rebbachisauridae, que quizás constituya una característica común de los diplodocoides, que aún necesita confirmación. Gracias a ellos, la cola creó una especie de látigo. Se podría usar para defender o hacer sonidos fuertes. Las apófisis espinosas se bifurcan, alcanzando una longitud de más de cuatro veces la altura que cuerpo. Probablemente hubo ligamentos entre las "horquillas". La bifurcación implicaba dividir el ligamento supraespinoso en dos partes.7 Guiado por esta característica de las vértebras, al animal se le dio el nombre genérico Dicraeosaurus que significa "lagarto bifurcado".
El primer fósil fue descrito por Werner Janensch en 1914. Se encontraron fósiles en la Formación Tendaguru en Tanzania y probablemente también de Kazdi en Zimbabwe. Cerca de estos fósiles, también se han encontrado restos de Giraffatitan y Kentrosaurus.
Dicraeosaurus proviene del famoso sitio de Tendaguru en Tanzania y fue encontrado durante las expediciones alemanas de 1909 a 1913. En Tendaguru se pueden distinguir tres capas diferentes en la que se producen los dinosaurios y con edades ligeramente diferentes, las margas de dinosaurios Inferior, Media y Superior. Los dos tipos de Dicraeosaurus por lo tanto no vivían juntos, se encontraron en diferentes capas, Dicraeosaurus hansemanni en la Inferior y Media entre el Oxfordiano y el Kimeridgiano y Dicraeosaurus sattleri en la Superior del Titoniano. El sitio en Tendaguru representa una zona de marea en la costa y por lo tanto, probablemente no sea el hábitat real de Dicraeosaurus, que se encuentra más en el interior.
En un principio se incluyó un Dicraeosaurus en la familia Diplodocidae, McIntosh consideró este punto de vista en 1990. Desde 1991 se lo ha colocado Dicraeosauridae, que es parte de la superfamilia Diplodocoide.18 En 2004, Harris y Dodson definieron el clado Flagellicaudata para incluir el último ancestro común de Dicraeosaurus y Diplodocus y todos sus descendientes. Se cree que Suuwassea emilieae, es un taxón hermano de todos los demás representantes del grupo. Se cree que la familia Dicraeosauridae es más avanzada en la evolución que la familia Diplodocidae.
A su vez, el clado que une el Dicraeosaurus y Brachytrachelopan se llamaba Dicraeosaurinae. Amargasaurus no pertenece a esta subfamilia. De acuerdo con el estudio de 1999 Salgado D. hansemanni en realidad tiene una relación muy estrecha con Brachytrachelopan, más cerca incluso que a D. sattleri, que parece ser el pariente más cercano a Amargasaurus. Esto indicaría que el género Dicraeosaurus que cubre estas dos especies es polifilético. Sin embargo, en el estudio de 2005 Rauhut confirmó el tipo de monofilia, así como su relación más cercana a Brachytrachelopan que a Amargasaurus. Además, Dicraeosaurus parece más basal que Amargasaurus, no es igual en las longitudes de las vértebras bifurcadas o el proceso alado. Sin embargo, se observa que es necesario llevar a cabo más estudios que confirmen la sistemática de este grupo. A su vez, el clado que une el Dicraeosaurus y Brachytrachelopan se llamaba Dicraeosaurinae. Amargasaurus no pertenece a esta subfamilia. De acuerdo con el estudio de 1999 Salgado D. hansemanni en realidad tiene una relación muy estrecha con Brachytrachelopan, más cerca incluso que a D. sattleri, que parece ser el pariente más cercano a Amargasaurus. Esto indicaría que el género Dicraeosaurus que cubre estas dos especies es polifilético. Sin embargo, en el estudio de 2005 Rauhut confirmó el tipo de monofilia, así como su relación más cercana a Brachytrachelopan que a Amargasaurus. Además, Dicraeosaurus parece más basal que Amargasaurus, no es igual en las longitudes de las vértebras bifurcadas o el proceso alado. Sin embargo, se observa que es necesario llevar a cabo más estudios que confirmen la sistemática de este grupo.
Sistemática[]
El siguiente cladograma sigue un análisis por Sereno et al. en 2007.
| Diplodocoidea |
|
Dicraeosaurus vivió durante el Jurásico superior, junto con los otros dinosaurios y parientes que vivieron durante ese tiempo. Era herbívoro, sin embargo, no compitió con otros dinosaurios para la comida. Los fósiles se han descubierto en las rocas de la Formación Tendaguru en Tanzania. Las rocas también brindan fósiles de Giraffatitan (incluido dentro del género Brachiosaurus por Janensch en 1914) y de Kentrosaurus. Esto presenta evidencia que estos eran los herbívoros con el que Dicraeosaurus vivió, así que no habría tenido que competir por el alimento ya que se alimentaban en distintos niveles.
En el Jurásico tardío, la vegetación tipo herbácea predominaba en las áreas correspondientes a las colinas de Tendaguru de hoy y la formación Morrison. Los árboles crecieron más abundantemente en las zonas costeras. Estas plantas eran alimento para saurópodos. Además de las dos especies en Dicraeosaurus Tendaguru encontrado restos de Giraffatitan brancai, Australodocus bohetii, Tendaguria tanzaniensis y Janenschia robusta. Las plantas altas, sin embargo, solo podían alimentar a algunos de los eusaurópodos. Incluso beneficiando a camarasáuridos, braquisáuridos y euhelopódidos, debido a la construcción adecuada del cuello. Además, gracias a sus dientes anchos, pudieron comer alimentos duros, como cícadas. Sin embargo, otros saurópodos como los dicreosáuridos y diplodócidos pastaban al nivel del suelo. Los diplocoideos por la morfología de sus dientes consumieron solo plantas blandas, y de acuerdo con algunas hipótesis, también filtraron el plancton. En general, sin embargo, las alturas de alimentación de los saurópodos individuales se superponen, no siendo las principales características distintivas de sus nichos ecológicos.
En la franquicia de Jurassic Park[]
Aparece en la dinopedia del sitio web extinto Jurassic Park Institute.